
Regenerasi merupakan suatu kemampuan dari neuron untuk memperbaiki dirinya sendiri, termasuk pembentukan kembali koneksi yang masih fungsional. Segera setelah suatu axon terputus, ujung proximal dari axon akan terbentuk pembengkakan atau suatu kerucut pertumbuhan (Thompson, 2006). Kerucut ini membentuk suatu psudopodia yang menyerupai kerucut pertumbuhan pada perkembangan saraf normal. Setiap kerucut pertumbuhan axon dapat membentuk banyak cabang dan tumbuh menjauhi dari lokasi potongan axon. Ketika cabang ini dapat melewati jaringan parut dan masuk pada ujung distal potongan saraf, maka regenerasi dengan pemulihan fungsi dapat terjadi.(Zochodne, 2008)
Proses perkembangan pertumbuhan saraf dan regenerasi saraf sepertinya serupa, akan tetapi terdapat perbedaan yang bermakna. Sel saraf embrionik dituntun oleh suatu faktor neurotropik, yang digunakan axon untuk inisiasi pertumbuhan kemudian mencapai targetnya. Sumber dari substansi neurotropik ini, yang dapat menstimulasi atau menginhibisi sinaps, termasuk kerucut pertumbuhan axon, dan matriks jaringan konektid atau sel stroma seperti sel glial dan fibroblast. Kemungkinan suatu axon tidak mencapai target perifer sangat dibatasi selama perkembangannya, dan hal ini diatur oleh pola genetik yang memberikan arah spesifik dari pertumbuhannya. (Thompson, 2006; Geuna, 2009)
Regenerasi, lebih terbatas pada inisiasi pertumbuhannya dalam keadaan yang tidak menguntungkan. Tidak seperti sel satelit pada otot, fibroblast pada kulit, dan hepatosit pada liver, saraf tidak memiliki cadangan sel untuk regenerasi. Sel Schwann yang mengalami degenerasi pada perifer dan faktor kemotaktik lainnya memberi sinyal adanya kerusakan axon pada motor neuron melalui transport retrograde dalam medulla spinal. Terdapat beberapa teori yang menjelaskan inisiasi regenerasi. Teori yang paling umum adalah adanya NGF dan komponen kemotaktik lainnya yang terdapat pada target perifer dan didalam selubung yang meliputi sel Schwann berkontribusi terhadap inisiasi dan keberlangsungan regenerasi saraf.(Zochodne, 2008)
Proses regenerasi idealnya mencakup beberapa tahapan penting yaitu sprouting awal, elongasi axon, pertumbuhan kaliber axon, remyelinasi dari kaliber axon, pemulihan saraf dengan axon yang matang dan kontak dengan jaringan target. Selama regenerasi axon mendapatkan kembali properti elektrofisiologisnya yang telah rusak sebelumnya. Pada akhirnya penting untuk diketahui bahwa telah terjadi pemulihan total dari saraf dengan melihat respon fungsional dari jaringan target.(Zochodne, 2008)
Dasar Seluler dan Biomolekuler Proses Regenerasi Saraf
Terdapat beberapa interaksi seluler yang menandai tahap dini regenerasi saraf setelah terjadi cedera saraf perifer. Elemen utama yang tejadi adalah pembentukan membran baru untuk sprouting, ekspresi gen yang berubah pada seluruh neuron, sintesis axonal, pergerakan konus pertumbuhan, dan perubahan fenotip pada sel Schwann yang memfasilitasi dan menuntun pertumbuhan axon baru. Semua elemen ini harus terkoordinasi untuk terjadinya regenerasi yang bermakna. (Zochodne, 2008)
Segera setelah cedera saraf, akan terjadi perubahan patofisiologis yang kompleks mencakup perubahan morfologi dan metabolik pada lokasi cedera. Perubahan ini juga terjadi pada sel tubuh saraf, segmen proksimal dan distal dari lokasi cedera, dan pada ujung distal dari muscle end plate atau reseptor sensorik. Perubahan pada lokasi cedera terjadi paling pertama. Organel dan metabolit berakumulasi pada bagian proksimal dan distal ujung saraf yang rusak, menyebabkan terjadinya pembengkakan saraf. (Jaweed, 2001)
Seluruh segmen distal lesi perlahan mengalami degenerasi. Dalam 6 jam setelah cedera, sel tubuh neuronal mengalami perubahan, tubuh Nissle dan neurofilamen terurai, akhirnya nukleus berpindah lokasi dari tengah sel menuju ke perifer sel untuk persiapan perubahan metabolik dari sintesis neurotransmitter menjadi produksi materi yang dibutuhkan untuk pertumbuhan dan elongasi axon. Sel harus mensintesis mRNA, lipid, dan protein terutama protein sitoskeletal seperti tubulin dan actin, neurofilamen, dan protein gap-associated. (Zochodne, 2008)
Sel Schwann yang berespon terhadap perubahan degenerasi axon dan myelin terlepas dari axon dan mulai bermultiplikasi, sehingga meningkatkan sintesis protein dalam selubung basal lamina. Proses ini disebut dengan degenerasi wallerian, sehingga menyebabkan axon mengalami degenerasi dibawah lesi. (Thompson, 2006) Sel Schwann berperan dalam memberi pola dan mendukung terjadinya regenerasi pada saraf perifer. Penelitian tekini mengindikasikan bahwa sel ini memiliki peran krusial dalam mempertahankan perkembangan axon. Ketergantungan motor axon terhadap keberadaan sel Schwann terlihat pada hewan dewasa, ketika terjadi sel Schwann patologis maka akan terjadi axon patologis, pada akhirnya menyebabkan kematian saraf. Penelitian lain menyebutkan bahwa pertumbuhan axon sebenarnya terjadi pada lamina basalis sel Schwann bukan pada permukaan sel Schwann. Tidak seperti kecepatan pertumbuhan yang normal, yakni 2-3 mm perhari, regenerasi axon yang tumbuh pada daerah luka lebih lambat yaitu sekitar 0,25 mm per hari. (Jaweed, 2001; Zochodne, 2008)
Pentingnya regenerasi axonal pada selubung sel Schwann yang liputi oleh basal lamina pada potongan distal menunjukkan terdapat beberapa tingkat regenerasi yang terlihat setelah saraf terperas dibandingkan transeksi saraf. Setelah cedera perasan, axon berada dalam keadaan parah, namun sel Schwann, lamina basalis sekitar, dan perineurium mempertahankan kontinuitas dalam lesi, sehingga dapat memfasilitasi regenerasi axon yang cedera. Berbeda ketika saraf terputus, maka kontinuitas dari mekanisme ini akan terganggu. (Jaweed, 2001)

Gambar 1. Gambaran peranan sel Schwan dalam reiinervasi neuromuscular junction
dijelaskan bahwa ketika terjadi degenerasi pada saraf maka akan terjadi perselubungan pada bagian distal lesi, degenerasi ini diikuti oleh aktivasi dan proliferasi sel Schwann pada bagian potongan distal. Lebih lanjut lagi, ”selubung endoneurial” yang terbentuk dari basal lamina disusun oleh sel Schwann dan perineurium yang meliputi fasikel axon tetap bertahan. Perselubungan ini tetap berada pada bagian distal dari saraf yang cedera dan menjadi saluran dimana axon akan tumbuh kembali setelah melewati lokasi lesi. Pentingnya selaput saraf yang tua dalam mengarahkan pertumbuhan axon telah dijelaskan dalam penelitian Nguyen et al. Pada penelitian ini terlihat pada saraf otot tikus yang telah dirusak, nampak axon yang terputus melacak jalur saraf lama untuk menuju ke otot dan kemudian berakhir pada serat otot yang sama sebelum saraf tersebut dirusak. Peranan dari selubung endoneurial dalam mengarahkan regenerasi axon pada lokasi sinaptik dalam otot telah disimpulkan dari beberapa eksperimen pencitraan sebelumnya (Jaweed, 2001) Pertumbuhan axon didahului oleh pembentukan kerucut pertumbuhan pada ujung potongan proksimal. Kerucut pertumbuhan ini disebut sebagai “tangan penjelajah” axon yang berregenerasi. Memiliki pengaruh terutama bagaimana regenerasi akan terjadi. Kerucut pertumbuhan terdiri dari filopodia berupa jari motil yang meliputi aktin filamen yang berasal dari lamellipodium yang memanjang dari area pusat dimana mikrotubul dan axoplasma berakhir. Komponen ini kaya akan retikulum endoplasmik, mikrotubulus, mikrofilamen, mitokondria yang besar, lisosom, dan struktur vakuola yang belum diketahui fungsinya. (Zochodne, 2008)
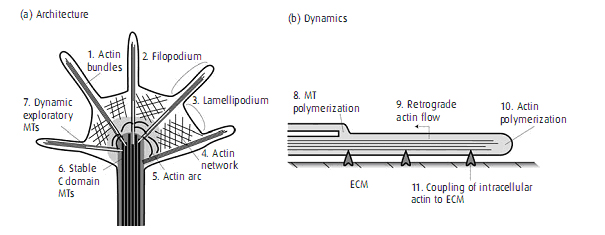
Gambar 2. Struktur dari kerucut pertumbuhan
Telah disimpulkan bahwa sebelum axon yang beregenerasi memanjang dari potongan proksimal, ujung dari kerucut pertumbuhan melekat pada kolagen yang meliputi potongan distal yang degeneratif. Filopodia pada kerucut pertumbuhan memanjang menuju potongan distal saraf setelah adanya peningkatan sitoskeletal protein yaitu aktin dan myosin pada tubuh sel. Kebanyakan kerucut pertumbuhan mencari ujung dari sel Schwann. Pergerakannya mendekati tempat dimana terdapat prekursor sel Schwann sehingga akhirnya kerucut pertumbuhan mendekati potongan distal dari saraf yang rusak. Dalam perjalanannya, dikatakan bahwa prekursor sel Schwann meliputi sekitar 80% dari permukaan kerucut pertumbuhan (Zochodne, 2008)
Percabangan regeneratif (sprout) dapat terbentuk baik pada ujung distal axon atau pada bagian proksimal. Beberapa jam setelah cedera, axon pada segmen proximal menghasilkan sprouting yang bertumbuh ke arah distal melalui selubung di dalam basal lamina. Gelombang sprouting pertama diikuti oleh gelombang berikutnya dalam 2 hari pertama. Sprouting awal dapat mengalami degenerasi sebelum fase sprouting definitif terjadi. Selang waktu hingga sprouting definitif terjadi disebut “initial delay“. Terdapat beberapa jenis sprouting yang diamati terjadi pada saraf sciatic tikus percobaan. Sprouting dapat berupa proyeksi langsung yaitu axon tunggal melewati celah antar segmen distal proksimal, dan proyeksi kolateral dimana axon tunggal membentuk percabangan secara paralel menuju potongan segmen lainnya. Karena saraf yang dikultur mengalami cedera pada saat pengambilan spesimen, maka mekanisme collateral sprouting sulit diamati in vitro. Hal ini disayangkan karena proses ini diperlukan untuk memahami pemulihan lesi saraf pada manusia. (Zochodne, 2008; Thompson, 2006 ) .
Degenerasi Wallerian pada potongan distal menyebabkan akumulasi material kolagenosa sekitar axon yang berdegenerasi. Sekitar 28 – 35 hari setelah cedera, kolagen endoneurial terkumpul pada bagian distal, semakin memberi tekanan pada axon yang beregenerasi sehingga mengecilkan diameter dan meningkatkan jumlah sel Schwann untuk tiap panjang axon yang mengalami regenerasi. Axon baru memiliki panjang internodal yang lebih pendek dan suplai vaskularisasi yang lebih sedikit dibanding sebelum mengalami kerusakan, bahkan tetap seperti itu hingga remyelinasi. Penelitian terkini mengindikasikan bahwa asetilkolin dan adenosine triphosphat (ATP) dilepaskan dari axon yang tumbuh dan bekerja melalui reseptor untuk merubah sel Schwann dari pertumbuhan myelin menjadi bermyelin. Neuregulin berperan penting untuk remyelinisasi axon yang berregenerasi. (Zochodne, 2008)
Tujuan akhir dari regenerasi saraf adalah untuk menciptakan kembali sinaps dengan organ target. Ujung saraf motorik tersambung dengan saraf di neuromuscular junction, dimana efek neurotropik bekerja pada otot. Ketika saraf motorik rusak, maka axon berregenerasi dan menciptakan sinaps lainnya, dapat pada lokasi sebelum cedera atau sekitarnya. Dalam waktu 2-3 minggu setelah denervasi, neuromuscular junction akan tercipta kembali dan konduksi saraf juga kembali normal.. (Zochodne, 2008)
Faktor Penentu Regenerasi Saraf
Pada saat terjadi degenerasi saraf perifer maka akan terjadi suatu lingkungan yang mendukung terjadinya regenerasi dan pertumbuhan kembali axon yang cedera. Faktor neurotrophik mendukung bertahannya neuron tertentu. Faktor neurotrophik yang dimaksud adalah neurotrophin (NGF, NPF, dan neurotrophin-4/5), insuline-like growth factor, ciliary neurotrophic factor, dan glial-cell derived growth factor (Zochodne, 2008).
Perubahan molekul yang terjadi pada bagian distal potongan saraf termasuk regulasi neurotrophin, adhesi molekul sel neural, sitokin, dan faktor lainnya beserta reseptornya. Kecepatan pertumbuhan axonal dan restorasi koneksi perifer diregulasi oleh beberapa faktor biokimia dan biofisik tertentu. Sebagai contoh, regenerasi sangat terkait dengan integritas selubung endoneurial. Ketika endoneurium tidak mengalami kerusakan, maka regenerasi axon berlangsung lebih mulus, bertumbuh dengan selubung yang utuh, dan pada akhirnya menemukan targetnya. Sebaliknya, jika endoneurial dalam keadaan rusak parah, maka perjalanan regenerasi akan lambat. Maka dari itu, pemulihan yang fungsional, yang berarti konduksi saraf dapat terjadi hingga ke organ target, kemungkinan lebih cepat terjadi pada cedera kompresi axon dibandingkan cedera traumatik atau transeksi. (Jaweed, 2001)
Neuron Growth Factor (NGF) disintesis pada jaringan target yang diinnervasi oleh neuron simpatetik atau sensorik dan mengalami transpor retrograde menuju ke neuron. NGF mempengaruhi navigasi neurite, morfologi kerucut pertumbuhan, dan elongasi axonal. Pemahaman mengenai biokimia dan faal NGF pada saraf perifer penting diketahui sebelum mengerti peranannya dalam mendukung terjadinya regenerasi saraf sensorik. NPF memiliki karakter yang serupa dengan NGF dimana mengikat pada substrat polikationik dalam kultur saraf dan pada lamina basalis dari sel Schwaan in vivo. Fibronectin, menyebabkan pemanjangan dari neurite pada kultur jaringan sehingga meningkatkan adhesi substrat atau matriks.(Geuna, 2009) Estrogen, testosteron, insulin, hormon adrenal dan thyroid, diketahui memiliki pengaruh terhadap regenerasi saraf dengan cara yang bervariasi. Sebagai contoh, estrogen dan insulin memfasilitasi pertumbuhan neurites pada kultur jaringan, dimana testosteron dan thyroid menstimulasi regenerasi nervus sciatic. Inhibitor protease seperti glial derive protease inhibitor dan leupeptin menghambat meluasnya degenerasi axon bagian distal, sehingga mempercepat terjadinya regenerasi. Fibroblast growth factor, nampaknya meningkatkan jumlah serat bermyelin pada tikus dan meningkatkan pertumbuhan neurite pada kultur sel PC-12. Efek FGF in vivo terkait dengan aktivasi sel Schwann dan memperkaya vaskularisasi. Inhibisi radikal bebas seperti catalase, rupanya mempercepat regenerasi karena mencegah terjadinya cedera oksidatif pada neuron yang rusak. (Jaweed, 2001)
Komponen lain yang memiliki efek positif adalah gangliosida cerebral. Glikolipid ini merupakan komponen dari membran plasma neuronal, disintesis pada neural soma, dan ditransport menuju perifer oleh transpor axonal yang cepat. Gangliosida yang diberikan pada hewan, menunjukkan peningkatan pembentukan sprouting saraf dan neuromuskular junction, sehingga menyempurnakan reinnervasi otot. (Geuna, 2009)
Regenerasi saraf perifer juga terkait oleh interaksi kompleks antara sel tubuh saraf, potongan proximal dan distal axon, dan faktor neurotrophic. Segera setelah cedera saraf, tubuh sel neuron pada medulla spinalis membengkak dan tubuh Nissl mulai mengalami degenerasi, dan nukleus bergerak menuju perifer untuk persiapan perubahan metabolisme dari sintesis neurotransmitter menjadi produksi materi yang dibutuhkan untuk pertumbuhan dan pemanjangan axon (Geuna, 2009).
Sel harus dapat mensintesis mRNA, lipid, dan protein yang baru, terutama protein sitoskeletal seperti tubulin, aktin, neurofilamen, dan protein gap-associated. Barrier antara darah dan saraf juga terbuka sepanjang saraf sehingga menyababkan invasi makrofag kemudian fagositosis aktif terjadi oleh sel ini. Faktor inhibisi terkait dengan myelin, dibuang dan sel Schwann secara progresif memanjang dan menyesuaikan basal lamina dari fibronektin dan laminin, sehingga terjadi lingkungan yang mendukung untuk axon yang berregenerasi dari potongan saraf bagian proksimal. (Zochodne, 2008)

{kind=link}